1. Signaali transduktsiooni mõiste ja signaali vastuvõtmine.
1.1. Signaali transduktsiooni mõiste.
1.2. Signaali toojad.
1.3. Kuidas signaal jõuab raku välispinnale?
1.4. Kuidas rakk võtab signaali vastu ? Retseptorid
1.4.1. Tsütoplasmaatilised retseptorid
1.4.2. Raku pinna retseptorid.
1.4.2.1. Ioonkanalit moodustavad retseptorid e. neurotransmitterite ionotroopsed retseptorid.
1.4.2.2. G-valkudega seotud retseptorid e. metabotroopsed retseptorid
1.4.2.3. Pinge-avatavad ioonkanalid.
1.4.2.4. Valgulisi faktoreid siduvad retseptorid.
1.1. Signaali transduktsiooni mõiste.
Elus rakk on pidevalt informatsiooni vahetuses teda ümbritseva keskkonnaga. Ta saab ellu jääda ainult siis, kui suudab võtta vastu keskkonnast tulevaid signaale ja neid oma tegevuses arvestada, vastata neile oma geneetilise informatsiooni baasil. Rakuni jõuab väljast väga palju igasuguseid signaale, mis muudavad mitmesuguseid biokeemilisi reaktsioone rakus. Üks raku välispinnale tulnud signaal käivitab raku sees keemiliste reaktsioonide rea, mida nimetatakse signaali ülekande protsessiks. Rahvusvahelises kirjanduses kasutatakse nimetust signaali transduktsioon. Signaali ülekanne e. transduktsioon on protsess kuidas rakk võtab vastu välissignaali, juhib selle läbi tsütoplasma tuuma ja organiseerib esmase vastuse. Eristatakse kolme etappi: 1 - kuidas rakk võtab vastu signaali, selles osaleb signaali tooja ja raku pinna retseptor; 2 - kuidas signaal liigub tsütoplasmas, siin toimuvad rea valkude aktiivsuse muutused ja madala molekulaarsete signaali vahendajate kontsentratsiooni järsud muutused; 3 - tuuma sisene etapp kujutab endast transkriptsiooni faktorite aktivatsiooni ja nn. esmase vastuse geenide ekspressioon.
Kõik signaali teed ei vii tuuma. Retseptori aktiveerimisele järgneb rida protsesse tsütoplasmas, mis reguleerivad valgu aktiivsust modifikatsioonide läbi. Valgu taset reguleeritakse ka translatsiooni tasemel, geeni ekspressiooni ei muudeta. Translatsiooni initsiatsioonil osalevad faktorid fosforüleeritakse retseptori aktivatsiooni järel ja see tõstab translatsiooni teatud mRNA-lt. Translatsiooni taseme regulatsioon toimub mRNA 5'- ja 3'UTR (mittetransleeritavate regioonide) kaudu. Ka ribosoomi valk S6 fosforüleerimisega muudetakse mõnede mRNA-de translatsiooni taset.
1.2. Signaali toojad.
Signaali toojateks on kõik välisfaktorid, mis mõjutavad rakku, alates kosmilisest kiirgusest ja lõpetades naaberrakkude kontaktidega. Oma olemuselt võib signaali kandjaid jagada füüsikalisteks, keemilisteks ja bioloogilisteks.
- Füüsikalised signaali kandjad.
1) Kiirgused. Praeguseks on uuritud valgus-signaali vastu võtmist ja sellest lähtuvaid muutusi rakus. Teiste kiirguste osa signaali tee aktiveerijana on vähem uuritud.
2) Elektriline pinge. Raku elutegevuse käigus muutub pidevalt ioonide kontsentratsioon raku välismembraani kummalgi pool, sellest tekib membraanis pinge. Pinge reguleerib membraansete ioonkanalite avamist.
B. Keemilised signaali kandjad.
Enamus praegu teada olevatest signaali kandjatest on keemilised ühendid. Keemilisi ühendeid on väga palju ja neid kui signaali kandjaid võib klassifitseerida kas molekulmassi, keemiliste omaduste, toime mehhanismi või veel teiste näitajate järgi. Siin on toodud mõned signaali kandvate ühentite rühmad:
1) Lõhna- ja maitseained, siia kuuluvad keskkonnast tulevad enamasti madalamolekulaarsed ühendid.
2) Neurotransmitterid, see nimetus on pärit neurobioloogiast ja tähistab aineid, mis osalevad närvi impulsi edasi andmisel. Siia kuuluvad näiteks klassikalised neurotransmitterid atsetüülkoliin, g -aminovõihape, glutamiinhape, glütsiin, asparagiinhape, türosiin ja tema derivaadid adrenaliin, noradrenaliin, dopamiin ning serotoniin. Signaali ülekande uurimisega tegeles pikka aega ainult neurobioloogia. Viimase ajal on signaali transduktsioon muutunud omaette molekulaarbioloogia haruks ja on selgunud, et mitmed neurotransmitteritena tuntud ained on signaali vahendajad ka teistes rakkudes. Eriti käib see aminohapete ja nende derivaatide kohta. On arvamusi, et kõik aminohapped või nende derivaadid on rakus kasutusel ka kui signaali vahendajad, kui transmitterid. Kirjanduses kasutatakse sageli madalamolekulaarsete signaalainete kohta ka termineid transmitter ja mediaator.
3) Eraldi rühmana tuuakse välja steroidhormoonid ja teised rasvades lahustuvad ühendid (östradiool e. estradiool, testosteroon, progesteroon, kortisool, A ja D vitamiinid jt), mille rakku mineku tee on erinev teistest, nad difundeeruvad läbi membraani. Siin on nii teiste rakkude poolt toodetud kui ka väliskeskkonnast tulevad ühendid.
4) Läbi raku membraani lähevad ka gaasilised ühendid nagu NO ja CO, mis on viimastel aastatel signaal-ainete hulka jõudnud. Nii teiste rakkude poolt toodetud kui väliskeskkonnast tulnud lämmastik- ja süsinikoksiid toimivad rakus signaali vahendajatena. NO ja CO on nii rakkude vahelised kui rakusisesed signaali vahendajad.
5) Peptiidid on sageli signaali kandjateks nii närvikoes kui mujal. Need on mõnest kuni mõne kümnest aminohappest koosnevad valgu jupid, mida rakud teatud tingimustel sekreteerivad ja millele teised rakud vastavad.
6) Valgulised hormoonid (insuliin) ja kasvufaktorid. See on suur ja üha kasvav signaali kandjate rühm. Rakkude kasvamine ja differentseerumine on reguleeritud sellega, milliseid kasvufaktoreid nad oma ümbrusest saavad. Mõni faktor paneb rakud kasvama, teine pidurdab kasvu.
C. Bioloogilised mõjurid.
Siia rühma võiks panna elusad signaali kandjad - viirused, bakterid, naaberrakud. Need bioloogilised mõjurid toimivad ka üle valkude. Kontaktid naaberrakkude vahel, rakkude ja viiruste ning bakterite vahel toimuvad mõlema poole pinna-valkude abil.
1.3. Kuidas signaal jõuab raku välispinnale?
Kiirguste puhul mingeid eri mehhanisme pole vaja, lõhna ja maitse ained ka difundeeruvad ise. Neurotransmitterite ja valguliste faktorite jaoks on kirjeldatud 4 signaali levimise teed:
1) Endokriinne - sel juhul signaalained kantakse rakkudeni mööda vereringet ja lümfisooni. Organismi mõned koed toodavad signaalaineid ja teised võtavad neid vastu ja reageerivad neile. Sellisel teel levivaid signaalaineid nimetatakse klassikaliselt hormoonideks. See tee on omane kõrgematele organismidele.
2) Sünaptiline - signaalained (neurotransmitterid) suunatakse kõrges kontsentratsioonis teise raku retseptorile. See on närvi impulsi ülekanne.
3) Parakriinne - rakk sekreteerib signaalainet ümbritsevasse keskkonda, naaberrakud võtavad selle vastu ja reageerivad sellele. See on kudede sisene signaali tee, koekultuuris kasvavate rakkude puhul ja ainuraksete organismide puhul.
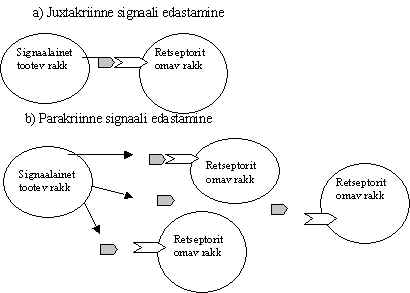
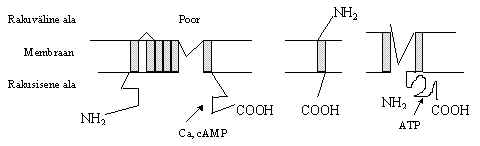
4) Juxtakriinne - naaberrakkude vaheline signaali edastamise tee, kus signaalmolekulid on membraaniga seotud. Seda võib vaadata ka kui parakriinse tee eri vormi, ühe raku adhesiooni valgud ja membraaniga seotud kasvufaktorid mõjuvad naaberrakule. Selle signaali edastamise viisi juures tekib küsimus kumb on signaali tooja ja kumb vastu võtja. Enamik neist faktoritest võib esineda ka lahustunud kujul, rakuväline ala on ära lõigatud ja see toimib kui parakriinne signaali kandja. Pildil 1 on kujutatud membraaniga seotud faktoriga ja vaba faktoriga signaali edasi andmine. Jxtakriinsete signaali kandjate hulka kuuluvad EGF-perekonna ja TNF-perekonna valgud. EGF on epidermiaalne kasvufaktor ja TNF on tuumor nekroosi faktor.
EGF-perekonna valgud on: EGF, TGF-a (transformeeriv kasvufaktor), betacellulin, amphiregulin, SDGF (Schwannoma-derived growth factor), HB-EGF (heparin-binding EGF-like growth factor), VGF (viral vaccinia growth factor), Lin-3 (C.elegans), Spiz (Drosophila), Gurken (Drosophila), Neuregulin. TNF-perkonna valgud on: TNF-a , lymphotoxin a , b , CD40-le ja CD27-le seostujad. Need faktorite perekonnad erinevad oma struktuurilt ja toimelt rakkudele.
Peale nende kahe rühma, on teada veel mitmeid juxtakriinselt signaali edastavaid faktoreid: c-kit retseptorile seostuja, CSF-1 (kolooniaid stimuleeriv faktor), CD21, Boss (Drosophila) - see on seostuja Sevenless retseptorile (Drosophila).
Pilt 1. Juxtakriinne ja parakriinne signaali edasi andmine.
1.4. Kuidas rakk võtab signaali vastu ? Retseptorid.
Juba 1870-ndatel aastal oli püstitatud hüpotees, et raku pinnal on punktid, kuhu seostuvad mitmesugused ühendid, mis rakule mõjuvad. Erinevad punktid on erineva affiinsusega teatud ainete suhtes. Raku pinnal signaali vastuvõttev punkt on retseptor. Retseptoriga seostuvat ühendit nimetatakse selle retseptori ligandiks. Retseptoreid on uuritud juba üle 100 aasta. Neid on klassifitseeritud ligandi sidumise järgi. Iga retseptor seob teatud affiinsusega signaali kandvat ühendit. Peale õige ligandi seovad retseptorid veel mõningaid tema analooge või ka hoopis teise struktuuriga ühendeid. Need on vastava retseptori agonistid või antagonistid. Agonistid seostuvad retseptoriga ja aktiveerivad selle, erinevalt õigest ligandist, seostuvad nad sageli pöördumatult ja aktiveerivad retseptori pikaks ajaks. Antagonistid seostuvad retseptoriga, ei aktiveeri seda ja ei lase ka õigel ligandil seostuda, inaktiveerivad retseptori. Nende ühenditega retseptorite aktiveerimist ja blokeerimist uurides jõuti järeldusele, et igale neurotransmitterile vastab mitu retseptorit, mis on tundlikud erinevatele agonistidele või antagonistidele.
Praegusel ajal võib retseptorid klassifitseerida ka muude tunnuste järgi. Lokalisatsiooni järgi jagatakse retseptorid raku pinna retseptoriteks ja tsütoplasmaatilisteks. Oma ehituse ja fuktsioneerimise järgi jagatakse neid rühmi veel mitmesse alarühma.
1.4.1. Tsütoplasmaatilised retseptorid.
Raku siseseid retseptoreid saavad kasutada signaalained, mis läbivad raku membraani. Näiteks gaasid ja rasvas lahustuvad ühendid, nagu steroidhormoonid, A ja D vitamiinid jt. Tsütoplasmaatiline valk võib peale väljast tulnud signaalaine sidumist algatada signaali tee, aktiveerida tsütoplasmaatilised ensüümid. Näiteks rakku tulnud NO ja CO seostuvad heemi-rühmaga ja aktiveerivad cGMP tootmise ja vastava signaali raja.
Kuid ligandiga seotud valk võib ka ise tuuma minna ja olla transkriptsiooni modulaator, seostudes teatud kindla järjestusega DNA-l. Seda DNA ala nimetatakse antud hormoonile vastavaks elemendiks. Näiteks, glükokortikoidi seostumise järel oma retseptorile, retseptor läheb tuuma ja seostub DNA osale, mida nimetatakse glükokortikoidi vastus-elemendiks e. respons-elemendiks. Selliseid valke, mis ligandiga seostumise järel lähevad tuuma ja moduleerivad transkriptsiooni, nimetatakse ka tuuma retseptoriteks, sest aktiveeritult nad lokaliseeruvad tuumas, omavad hormooni sidumise kohta ja DNA sidumise kohta, mis tagab retseptori seostumise hormoonile vastava alaga promootoris. Tuuma retseptorite perekonda kuuluvad steroid- ja türoidhormooni ning vitamiin A ja D retseptorid. Tuuma retseptoreid grupeeritakse ka mitmesse alarühma vastavalt dimeriseerumise ja DNA-ga sidumise omapärale.
1.4.2. Raku pinna retseptorid.
Kui sai võimalikuks retseptori valkude geenide kloneerimine, on nende uurimine tormiliselt arenenud. Esimestena kloneeriti 1983. aastal rodopsiini cDNA ja 1986. aastal nikotiin-atsetüülkoliini retseptori ühe subühiku cDNA. Atsetüülkoliini retseptorit on põhjalikult uuritud, sest teda saadi eraldada suures koguses elektri angerja elektri organist, seal olevat kuni 20% elektri organi valgust retseptori valk.
Raku pinna retseptorid jagatakse nende ehituse põhjal mitmesse rühma.
1.4.2.1. Ioonkanalit moodustavad retseptorid e. neurotransmitterite ionotroopsed retseptorid.
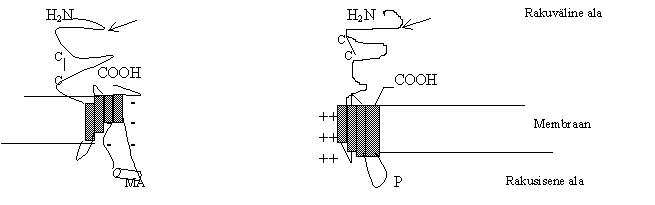
Siia rühma kuuluvad neurotransmitteritega aktiveeritavad ioonkanalid. Need on viiest alaühikust koosnevad s.t. pentameersed kompleksid raku membraanis, mis moodustavad kanali. Kanal laseb läbi katioone või anioone, kui ta on aktiveeritud. Tavaliselt koosneb kanal mitmest erinevast alaühikust e. subühikust. Erinevais kudedes koosneb kanal erinevatest alaühiku valkudest ja ka samas koes ei ole kõik kanalid kokku pandud täpselt ühesugustest valkudest. Näiteks, nikotiin-atsetüülkoliini (nAch) retseptor närvikoes koosneb kahest a ja kolmest b subühikist - a 2 b 3 , kusjuures on teada 7 erinevat a subühiku valku ja 3 erinevat b subühiku valku. Lihaskoes on nAch retseptor kokku pandud neljast erinevast subühikust: a 2b g d ja ka neid subühiku valke on mitu. Nii et erinevaid retseptoreid saab kokku panna üsna suure arvu. Pildil 2 on ionotroopsete retseptorite subühikute näited.
Pilt 2. Ionotroopsete retseptorite subühiku skeemid.
a) nAch retseptori a subühik ja b) GABA retseptori b subühik.
a) nAch retseptori a subühik omab 4 transmembraanset ala, N- ja COOH-ots on rakust väljas, tsütoplasmas on suur luup 3. ja 4. membraanse ala vahel. nAch retseptor on katiooni kanalit moodustav retseptor, laseb läbi K+, Na+ ja Ca2+ ioone, kanalis on negatiivsete laengute kogum, mis püüab katioone. Ekstratsellulaarses osas on N-glükosüleerimise kohad (nool) ja tsüsteiini sillad (C-C). Tsütoplasmaatilises osas võib olla veel spetsiifilisi piirkondi: sellel pildil MA tähistab valgu a -spiraali, mille üks pool on polaarne ja teine hüdrofoobne. Retseptorit võib moduleerida nii seest kui väljast poolt. Ligand atsetüülkoliin seostub pentameerse kompleksiga.
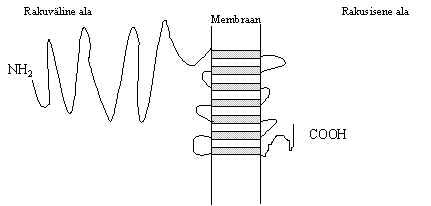
b) GABA-retseptor - g -aminovõihappe retseptori b -subühik. See on anioone läbilaskev kanal (Cl-, HCO3-), kanalis on positiivsete laengute kogum. Ka sel subühikul on 4 transmembraanset ala ja retseptor ise on pentameer a , b ja g subühikutest, igat subühikut on kirjeldatud mitu vormi. Väljas on glükosüleerimise kohad ja C-C sillad ning tsütoplasmaatilises osas on fosforüleerimise kohad (P).
Ionotroopsetest retseptoritest on üsna hästi uuritud ka glutamaadi ionotroopne retseptor, mis on analoogne oma ehituselt atsetüülkoliini ionotroopse retseptoriga, moodustab ka katiooni kanali. N- ja C-ots on väljas, tsütoplasmas on suurem aas 3. ja 4. transmembraanse ala vahel, sealt saab kanali avamist-sulgemist mõjutada. Ioonkanalit moodustavad glutamaadi retseptorid on jagatud kahte alarühma. On kas AMPA-kainaat-retseptorid, s.t. avatavad a -amino-3-hüdroksü-5- metüül-isooxsasool propioonhappega ja kainaadiga, või NMDA-retseptorid, avatavad N-metüül-D-aspartaadiga. Nii AMPA- kui NMDA-retseptorid on pentameersed. AMPA-retseptoril on teada 6 erinevat subühiku valku. Erinevused on sageli ainult üksikutes aminohapetes, kui muutused on transmembraanses alas, muutub kanali poolt läbilastavate ioonide koosseis: Ca-iooni ei lase läbi kõik AMPA retseptor-kanalid. Sageli saadakse üksikute aminohapete asendamine posttranskriptsiooniliselt mRNA editeerimise protsessis, s.t. muudetakse mRNA-s mõned nukleotiidid. Nii saadakse ühelt geenilt mitu erinevat subühiku valku. See on jälle üks retseptorite heterogeensuse tekitamise viise. NMDA-resteptor vajab ekstratsellulaarses osas aktiveerimiseks ka glütsiini, ta on blokeeritav Mg-ja Zn-ioonidega. Ka siin on mitut liiki subühikuid, retseptor võib olla homo- ja heteropentameer. Transmembraansed alad on lähedased AMPA-retseptorile. Tsütoplasmaatilises osas on fosforüleerimise kohad Ca/kalmoduliini kinaasile ja PKC-le.
1.4.2.2. G-valkudega seotud retseptorid e. metabotroopsed retseptorid.
See on suur erinevate ligandide poolt aktiveeritavate retseptorite rühm. Need retseptorid koosnevad ühest polüpeptiidist, mis läbib membraani 7 korda. Sageli kasutatakse ka nimetust - seitsme hüdrofoobse alaga polüpeptiid-retseptorid. Nimetus metabotroopne retseptor tähendab, et retseptori aktiveerimisele järgneb rakus mingi metabolismi käivitamine. Nimetus G-valguga seotud retseptor näitab, et need retseptorid kasutavad signaali edasi andmiseks ja metabolismi käivitamiseks G-valke. Kirjanduses kasutatakse kõiki kolme nime varianti. Seda tüüpi retseptor aktiveerib rakus erinevaid protsesse. Siin käivitatakse signaali tee mingite keemiliste reaktsioonide reana, ioone läbi retseptori sisse ei lähe. Retseptor võib küll reguleerida ka ioonkanaleid, aga need asuvad eemal ja aktiveeritakse vahendajate abil. Plidil 3 on seda tüüpi retseptori näiteks toodud glutamaadi retseptor.
Pilt 3. Glutamaadi metabotroopne retseptor.
Eespool oli jutt glutamaadi ionotroopsetest retseptoritest, glutamaadi aktiveeritav on ka teist tüüpi retseptor, metabotroopne retseptor. See on üks polüpeptiid, läbib 7x membraani, N-ots on väljas ja C-ots on rakus. Glutamaat seostub retseptoriga transmembraanse ala kaudu. Glutamaadi jaoks on teada 6 erinevat seda tüüpi retseptori valku. Erinevates kudedes ekspresseeruvad erinevad isovormid. Mõned glutamaadi retseptorid aktiveerivad cAMP lagundava diesteraasi, teised tõstavad inositooltrifosfaadi ja Ca-ioonide taset.
Metabotroopsete retseptorite rühma kuuluvad valgustundlik retseptor, lõhna ja maitse retseptorid, paljude neurotransmitterite ja lühikeste peptiidide retseptorid ning ka mõned valke siduvad retseptorid. Seitsme transmembraanse alaga on ka opioidide (oopiumi alkaloidid) retseptorid. Siia rühma kuuluvad ka muskariin-atsetüülkoliini retseptorid (mAch), nagu paljude neurotransmitterite nii on ka atsetüülkoliini jaoks mitu erinevat retseptorit: ionotroopne nAchR ja metabotroopne mAchR. Mõlemat tüüpi retseptorid on ka GABA jaoks. Üks moodustab ioonkanali, teine aktiveerib rakus metaboolse ahela.
Nii et ühele neurotransmitterile vastab mitu erinevat tüüpi retseptorit. Erinevad retseptorid osalevad erineva vastuse kujundamisel. Näiteks, glutamaadi retseptoritest ionotroopsed AMPA- kainaadi retseptorid annavad kiire vastuse - millisekundid, NMDA-retseptorid - 10-100 msek. ja metabotroopse retseptori vastusele kulub sageli tunde. Erinevatel retseptoritel on ka erinev tundlikkus blokaatorite ja aktivaatorite suhtes ning erinev affiinsus ligandi suhtes. Retseptorite paljusus võimaldab paremini reageerida keskkonna muutustele.
Nii ionotroopsed kui metabotroopsd retseptorid reguleerivad ioonkanaleid. Kanalid on olulised raku ja keskkonna vahelises suhtlemises. Vastavad ioonide pumbad, K+/Na+-ATPaas ja Ca2+-ATPaas, hoolitsevad selle eest, et raku sees oleks madal Na+ ja Ca2+ tase ja kõrge K+ tase. Ioonkanalites liiguvad ioonid vastavalt kontsentratsioonile, suuremalt kontsentratsioonilt madalamale. Neurotransmitterid reguleerivad kanalite avamist ja sulgemist, on väljast ja seest reguleeritavad kanalid (Tabel 1). Väljast avatavad ioonkanalid on ionotroopsed retseptorid: GABA ja glütsiini ionotroopsed retseptorid lasevad läbi anioone, nikotiiniga aktiveeritav Ach retseptor, glutamaadi AMPA- ja NMDA-retseptorid aga katioone. Subühikul on 4 transmembraanset ala.
Seest poolt reguleeritavad ioonkanalid aktiveeritakse mitmesuguste rakusiseste signaali vahendajate poolt (cAMP, cGMP, Ca-ioonid jt.). Viimaste kontsentratsiooni rakus muudavad metabotroopsete retseptorite poolt käivitatud reaktsioonid. Nii et raku seest reguleeritavad kanalid on metabotroopsete retseptorite kontrolli all. Seest avatavad või suletavad kanalid on teistsuguse ehitusega kui väljast avatavad: subühikul on 6 transmembraanset ala. Mõnede ioonkanalite omadused on toodud tabelis 1. Seal on kaks rühma transmitter-reguleeritavaid kanaleid, väljast või seest reguleeritavad.
Tabel 1.Transmitter-reguleeritavad ioonkanalid.
Ligand Läbivad ioonid Transmembraanseid alasid
subühikusA: väljast aktiveeritavad
GABA Cl-, HCO3- 4 glütsiin Cl-, HCO3- 4 nikotiinAch Na+, K+, Ca2+ 4 glutamaat/NMDA Na+, K+, Ca2+ 4 glutamaat/AMPA Na+, K+, (Ca2+) 4 serotonin (5-HT3) Na+, K+ 4 ATP (P2x kanali avamine) Ca2+, Na+, Mg2+ 4 B. raku seest reguleeritavad
cGMP (fotoretseptorite aktiveerimisel) Na+, K+ 6 cAMP, cGMP (lõhna rets. akt.) Na+, K+ 6 ATP (kanali sulgemine) K+ 6 IP3 (Ca hoidlates) Ca2+ (6)
1.4.2.3. Pinge-avatavad ioonkanalid.
Peale neurotransmitteritega avatavate ioonkanalite on veel pinge-avatavad ioonkanalid. Raku välismembraani kummalgi pool olevate ioonide kontsentratsiooni muutus tekitab membraanis pinge. Teatud tasemest alates on pinge võimeline avama spetsiifilisi ioonkanaleid. Neid on nii katiooni kanaleid kui ka aniooni kanaleid. Neile kanalitele on iseloomulik kõrge selektiivsus, iga kanal laseb läbi ainult ühte tüüpi ioone. Ka seda ühte tüüpi ioone läbilaskvat kanalit on kirjeldatud mitu erinevat vormi, mis on avatavad erinevate membraani pingetega.
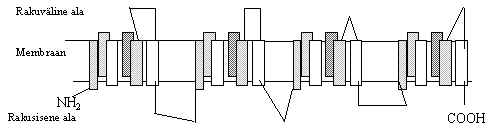
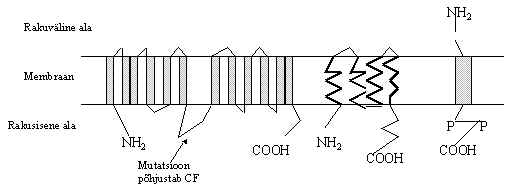
Pildil 4 on toodud mõned näited valkudest, mis moodustava ioonkanaleid Na, K või Cl ioonide jaoks. Oma ehituselt võivad nad olla üsna erinevad: on suuri palju kordi membraani läbivaid valke, nagu Na+-kanal. Samal ajal võib ioonkanali moodustada ka väike üks kord membraani läbiv valk. Kõigi nende valkude kohta pole teada mehhanism, kuidas nad kanali moodustavad. Näiteks, pildil toodud väikeste K+-kanali valkude puhul on teada, et sellise valgu cDNA rakku viimine tekitab seal K+-kanali, võib-olla osaleb seal lisaks sellele valgule ka teisi.
Na+kanal - üks suur a subühik 230-370 kDa, mõnes koes lisandub veel üks väike subühik 33-36 kDa. Suures subühikus on 4 suurt hüdrofoobset blokki, 3. ja 4. bloki vahel on regulatsiooniks oluline ala, mutatsioonid teatud kohtades põhjustavad haigusi. On mitu subühiku valgu geeni ja esineb ka erinevat splaisingut, avaldub koespetsiifiliselt.
K+ kanalid, nimetused on pärit Drosophila geenidest, kuid analoogseid valke on leitud ka teistes liikides: 1) Shaker-like kanalid - tetrameerid, subühik sisaldab kuni 6 transmembraanset ala, poori moodustav ala P on membraanis b-struktuuris, regulatsiooni alad on N- ja C-otsas.
2) Mink-kanalid - väike transmembraanne valk. 3) ATP-tundlik kanal, suletakse kõrge rakusisese ATP kontsentratsiooni korral.
Cl- kanalid: Cl-kanalid reguleerivad raku mahtu ja vee liikumist. Pildil esimene on CFTR (cystic fibrosis transmembran conductance regulator) selle Cl-kanali valgus ühe aminohappe asendamine põhjustab haigust - tsüstilist fibroosi. Haigetel on rikutud kloriidide ja vee liikumine läbi raku membraani, koe vedelikud on väga viskoossed. Teised näited on väiksematest transmembraansetest valkudest, mis moodustavad ka Cl-kanaleid, täpne kanali ehitus pole teada. Vaatamata kanalite paljususele, juba ühe kanali rike põhjustab surmava haiguse.
Siin toodud näited ei kirjelda kõiki kanalit moodustavaid valke, kuid juba nende järgi võib öelda, et kanali valke on mitmesuguseid ja ühine omadus on ainult see, nad on membraansed valgud.
Pilt 4. Ioonkanalid.
Na+ kanali a subühik
K+ kanalid 1) 2) 3)
Cl- kanalid 1) 2) 3)
1.4.2.4. Valgulisi faktoreid siduvad retseptorid.
Suure rühma moodustavad retseptorid, mis seovad valgulisi faktoreid ja on nende poolt aktiveeritavad. Nad on üks kord membraani läbivad valgud, millel on väljas ligandi siduv ala ja tsütoplasmas signaali edastav ala. Neid võib jagada mitmesse alarühma vastavalt sellele, kas nad omavad ise mingit ensümaatilist aktiivsust või ei oma, ja millist ensümaatilist aktiivsust nad omavad, ja kuidas signaali edasi antakse. Kirjanduses kasutatakse sageli nende kohta ka nimetust ensüümiga seotud retseptor, see pole eriti täpne nimi, sest G-valkudega seotud retseptor on ka ensüümiga seotud.
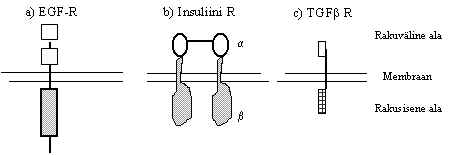
Pildil 5 on toodud mõned näited seda tüüpi retseptoritest.
Pilt 5. Valgulisi faktoreid siduvad retseptorid.
Kasvufaktorite retseptorid on sageli türosiinkinaasid või seriin/treoniinkinaasid
d) Tsütokiinide retseptorid ei oma mingit ensümaatilist aktiivsust:
e) Mitmest subühikust koosnevad ka T ja B rakkude antigeeni retseptorid:
a) Türosiinkinaassed retseptorid (TKR) - retseptori valk on üks peptiid, omab ühte transmembraanset ala, mille N-ots on väljas ja C-ots raku sees. Kui ekstratsellulaarsele osale seostub ligand, aktiveeritakse raku siseses osas türosiinkinaasne aktiivsus, s.t. võime fosforüleerida türosiini OH-rühmast. Retseptori ahelad dimeriseeruvad ja võivad teineteist fosforüleerida. Siia tüüpi kuuluvad PDGF (trombotsüütidest pärinev kasvu faktor), NGF (närvi kasvu faktor), EGF (epidermiaalne GF), FGF (fibroblastide GF) jt. kasvufaktorite retseptorid. Ka neid retseptoreid on palju ja erineva affiinsusega, ekstratsellulaarne ala võib üsna suuresti varieeruda. Näiteks FGF retseptorite varieeruvus on suur, läheneb immunoglobuliinide varieeruvusele.
b) Insuliini retseptor on ka türosiinkinaas, kuid natuke teise ehitusega: koosneb neljast subühikust, väljas on a-subühik, transmembraanne ja tsütoplasmaatiline ala on b-subühikust, kahe subühikuline retseptori ahel on veel dimeriseerunud. Kui teised TK-retseptorid dimeriseeruvad aktivatsiooni käigus, siis insuliini retseptor on pidevalt dimeer.
c) Mõned retseptorid omavad rakusiseses alas seriin/treoniin-kinaasset aktiivsust, nad fosforüleerivad seriini ja treoniini OH-rühmast. Sellised on näiteks TGFb (transformeeriv kasvu faktor) retseptorid.
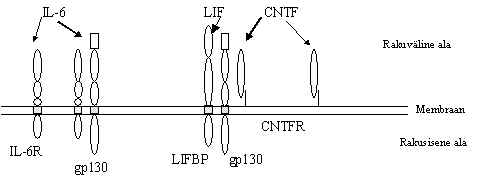
d) Tsütokiinide retseptorid. See retseptorite rühm ei oma rakusiseses alas mingit ensümaatilist aktiivsust, mõnedel retseptoritel puudub rakusisene ala hoopis. Need retseptorid vajavad signaali rakku viimiseks abivalku, mis ei seo ligandi aga seostub ligandi siduva valguga ja annab signaali edasi. Mitmele interleukiini retseptorile on abivalguks glükoproteiin gp130. Seda kasutavad IL-6 (interleukiin-6), LIF (leukeemia inhibiitor-fakor), oncostatin M (inimese melanoomi rakkude kasvu inhibiitor) ja CNTF (ciliary neurotrofic factor) retseptorid. Viimane neist ei oma üldse rakusisest osa ja ta vajab peale gp130 ka LIFBP (leukeemia inhibiitor faktorit siduvat proteiini). Signaali edastav retseptori kompleks koosneb mitmest erinevast valgust. Need kompleksid ei oma ise mingit ensümaatilist aktiivsust, nad aktiveerivad tsütoplasmaatilisi türosiinkinaase. Tsütokiinide retseptorite rühm on üsna heterogeenne, mõned tsütokiinid (näit. IL-8) kasutavad seitsme hüdrofoobse alaga retseptorit. Ligandi seostumine aktiveerib retseptori ja edasi G-valgu. On teada, et kui väiksed neurotransmitterid seostuvad seitse korda membraani läbiva retseptoriga just transmembraanse ala kaudu siis valgulised faktorid seostuvad ekstratsellulaarse ala kaudu.
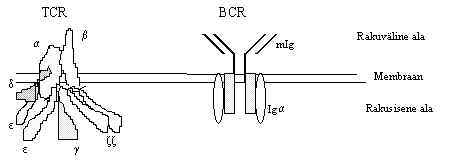
e) Suured mitme subühikulised kompleksid on ka T ja B rakkude antigeeni siduvad retseptorid.
TCR - väljas on antigeeni tundvad a ja b -subühikud, millel tsütoplasmaatilist osa peaaegu ei ole, nad seostuvad membraanis teiste subühikutega (2 tseeta (z) alaühikut seostuvad b-subühikuga; 2 epsilon(e)-, gamma(g)- ja delta(d)-alaühikut on seotud a-ga), need kompleksid seostuvad tsütoplasmaatiliste proteiin-türosiin-kinaasidega, mis fosforüleerivad raku siseseid subühikuid.
BCR - väljas on antigeeni tundvad immunoglobuliinid (mIg - membraansed Ig), sisemised subühikud seostuvad türosiinkinaasidega ja annavad signaali edasi. Iga ja Igb on heterodimeeridena seostunud membraansete Ig-dega, kinaasset aktiivsust ei oma ükski subühik, nad aktiveerivad tsütoplasmaatilisi kinaase.
Kõigile kirjeldatud retseptoritele on iseloomulik:
1) Ligandi spetsiifilisus, 2) Ligandiga aktiveeritavus, 3) Signaali edasi andmine, 4) Suur heterogeensus. Viimane tagab rakulise vastuse väga mitmesugustes tingimustes ja mitmesuguse vastuse.
Peale nende eespool toodud retseptorite on raku pinnal veel palju pinna valke ja adhesiooni molekule, mis käituvad retseptoritena. Ka viiruste rakku tungimiseks on vaja raku pinnal teatud retseptorite olemasolu. Mitmete viiruste jaoks on need juba teada, sageli on nad integriini taolised pinnavalgud. Sageli on iseloomulik kahe (või rohkema) astmeline seostumine. Esiteks, kiire viiruse sidumine ühe retseptoriga, see on nõrk side ja see võib toimuda üsna madalal temperatuuril, see on pöörduv protsess ja siin võib viiruse raku pinnalt veel maha pesta. Teiseks, aeglane etapp, viirus seostub järgmise retseptoriga või mitme retseptoriga, mis on esimese seostumise lähedal, või mis on nihkunud sinna lähedale esimese seostumise tõttu. See seostumine on tugevam ja sellele järgneb viiruse rakku minek kas endotsütoosi teel või siis nukleiinhappe rakku saatmine, viiruse nukleiinhappe rakku minek põhjustab nakkumise. Selleks, et viirus rakku nakataks on vaja mõlemat tüüpi retseptorit ja viiruse seostumist nendega. Näiteks, HIV seostub CD4 retseptoriga, kuid see pole piisav viiruse rakku minekuks. Nakatumiseks on vaja ka teist retseptorit (sekundaarne retseptor). HIV-1 ja HIV-2 kasutavad erinevaid sekundaarseid retseptoreid. Viiruste jaoks ei ole ainult nendele spetsiifilisi retseptorit, nad kasutavad mingit olemas olevat raku pinna retseptorit. Viiruseid võivad rakku transportida ka mõned aminohapete või fosfaadi transport-süsteemid. Leukeemia viiruse MLV-E retseptor on üks katioonseid aminohappeid transportiv valk (CAT), mis viib normaalselt rakku arginiini, lüsiini ja ornitiini. Viiruse seostumine retseptoriga takistab retseptori normaalset tööd ja rakku tuuakse viirus. GLVR1(gibbon ape leukemia virus receptor 1) on retseptor mitme leukeemia viiruse jaoks ja samal ajal anorgaanilise fosfaadi transportija. Mitmete madalamolekulaarsete ühendite transport rakku on seotud ioonide liikumisega läbi membraani. Viiruste retseptorite kohta on teada, et CAT töötab sõltumatult Na-ioonidest, GLVR1 on sõltuv Na-ioonidest ja võib olla inhibeeritud, kui Na on asendatud K- või Li-ioonidega.